
【一区家族】亚麻UGT家族全基因组鉴定及LuUGT175调控木质素生物合成的功能验证
一、文章基本信息
发表时间 :2022年10月4日
第一单位 :南通大学生命科学学院(江苏南通)
通讯作者 :孙健( sunjian8416@163.com )、苏建光( jianguangsu@163.com )
文章题目 :Comprehensive analysis of the UDP‑glycosyltransferase gene family in flax [ Linum usitatissimum L.] and functional verification of the role of LuUGT175 in the regulation of lignin biosynthesis
发表期刊 :Industrial Crops & Products
DOI:

二、摘要
背景:亚麻是全球重要的纤维作物,但纤维中木质素含量高会导致质地硬脆、可纺性差。UDP-糖基转移酶(UGTs)参与植物次生代谢,其糖基化修饰可影响木质素单体转运与聚合。
方法:本研究利用保守PSPG基序在亚麻基因组中鉴定出190个UGT基因(LuUGTs),进行系统进化、基因结构、表达模式分析;并通过CRISPR/Cas9敲除、组织化学染色、双荧光素酶报告系统等手段验证LuUGT175的功能。
结果:LuUGTs分为16个进化组,扩增主要依赖全基因组/片段重复;LuUGT175在茎与纤维中高表达,敲除后植株矮化、茎木质部染色加深且纤维层出现异位木质化;LuNAC61被鉴定为其上游负调控转录因子,可直接结合LuUGT175启动子。
结论:LuUGT175通过糖基化调控木质素单体稳态,进而影响亚麻细胞壁木质化,为低木质素、高品质亚麻分子育种提供理论依据和基因资源。
三、主要内容
图1
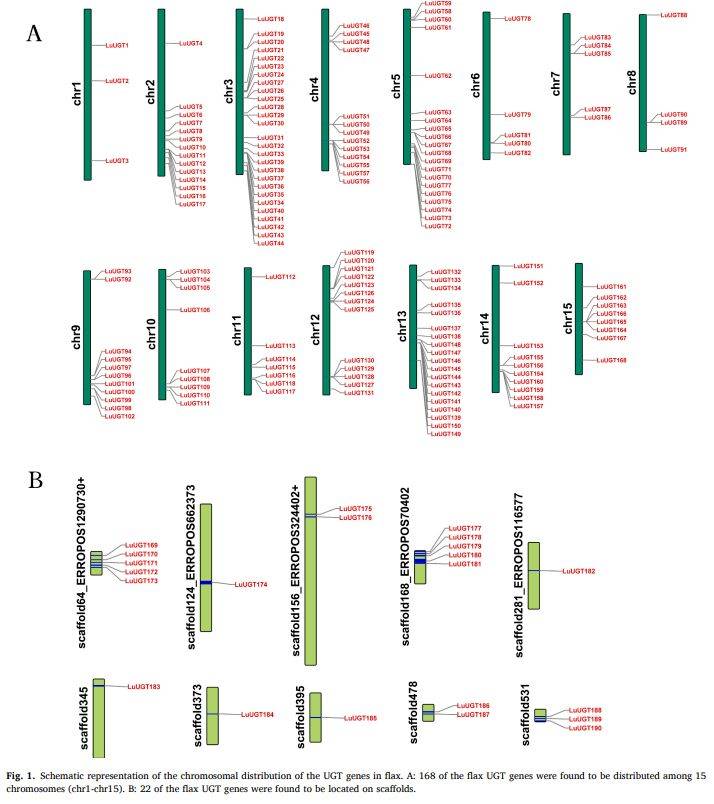
段落内容 :作者利用HMM与PSPG盒双重筛选,在亚麻全基因组中获得190条LuUGT候选序列。为展示其染色体分布格局,将168条定位于15条染色体,22条仍留在scaffold。图1A显示Chr3基因最密集(27个),Chr1最少(仅3个),呈现明显的不均衡分布;图1B给出22个scaffold-level基因的具体位置。该分布模式提示亚麻在进化中经历了大片段重复与染色体重排事件,为后续分析基因扩增机制提供线索。
展开全文
图例说明 :图1A为15条染色体上LuUGT基因位置示意,染色体编号位于底部,每条染色体右侧标注基因编号;图1B列出22个位于scaffold的LuUGT编号及对应scaffold名称。

图2
段落内容 :为解析LuUGT家族的进化关系,作者将190条亚麻UGT与19条拟南芥、3条玉米UGT共同构建邻接法系统树(bootstrap=1000)。结果将212条序列划分为17个亚组(A–Q),其中亚麻占据16组(Q组缺失)。最大的L组含29个成员,A、D、E、G组也都超过20个,合计占家族69.5%,说明这些分支在双子叶植物扩张最迅速。树形还显示LuUGT175与拟南芥AtUGT72B1处于同一E组小支,为后续功能研究提供直系同源依据。
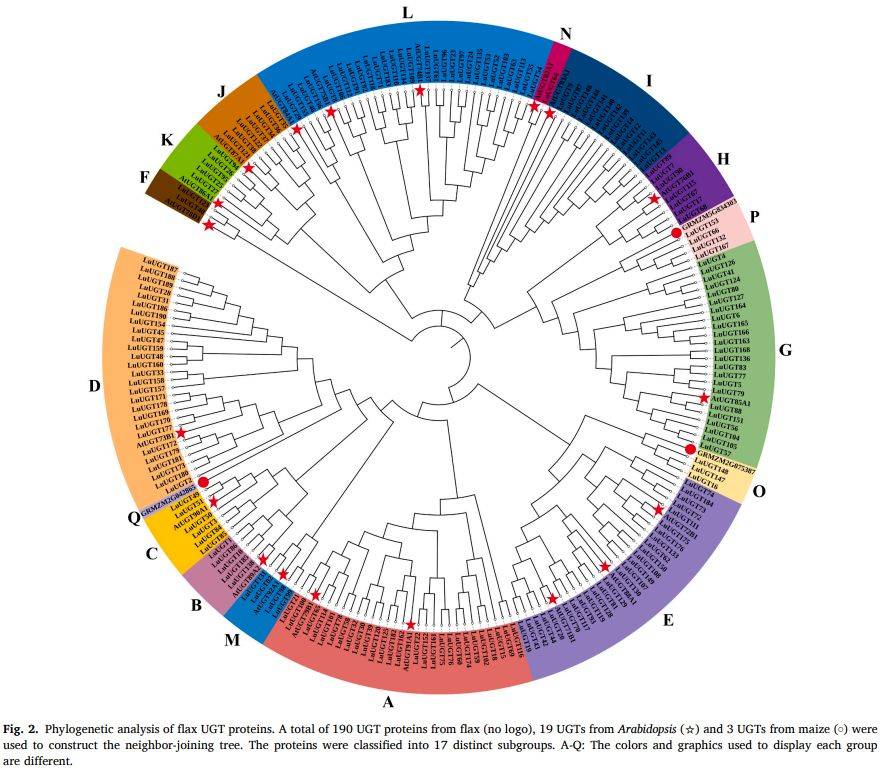
图例说明 :图2采用彩色扇形展示17个亚组,亚麻序列无图标,拟南芥标星,玉米标圆;分支长度代表氨基酸差异,bootstrap值>50%标注于节点。

图3
段落内容 :利用MEME与CDD对190条LuUGT进行保守元件解析,发现所有成员均含 motif1(UDGT催化域),171条同时含motif1-3;基因结构方面,60条无内含子,其余1–14个内含子不等,外显子数2–3个最常见。图3将系统树、motif构成、结构域及外显子-内含子组织四图并联,可直观看出同一亚组内基因结构高度相似,不同组间则差异显著,支持系统分组可靠性,也为后续选择E组LuUGT175做功能研究提供结构依据。
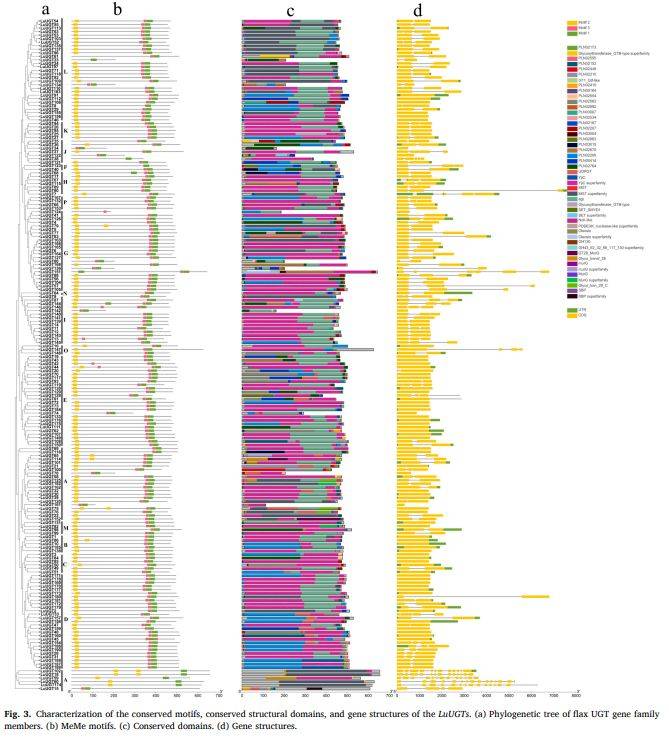
图例说明 :图3a为LuUGT进化树;b为motif1-3彩色方块图;c为NCBI-CDD预测结构域;d为外显子(绿色框)与内含子(灰线)排列,UTR以蓝色框表示。

图4
段落内容 :通过TBtools共线性模块,作者在全基因组范围鉴定到21对全基因组重复(WGD)与25对片段重复,占家族扩增主体;图中红线连接重复对,彩色背景块显示染色体间共线区。该结果首次从基因组水平证实WGD与局部重复是LuUGT家族扩张的主要动力,也解释了为何多数基因呈簇状分布。结合Ks值(未图示),大部分重复事件与双子叶古代γ三倍化及亚麻特异全基因组复制吻合。
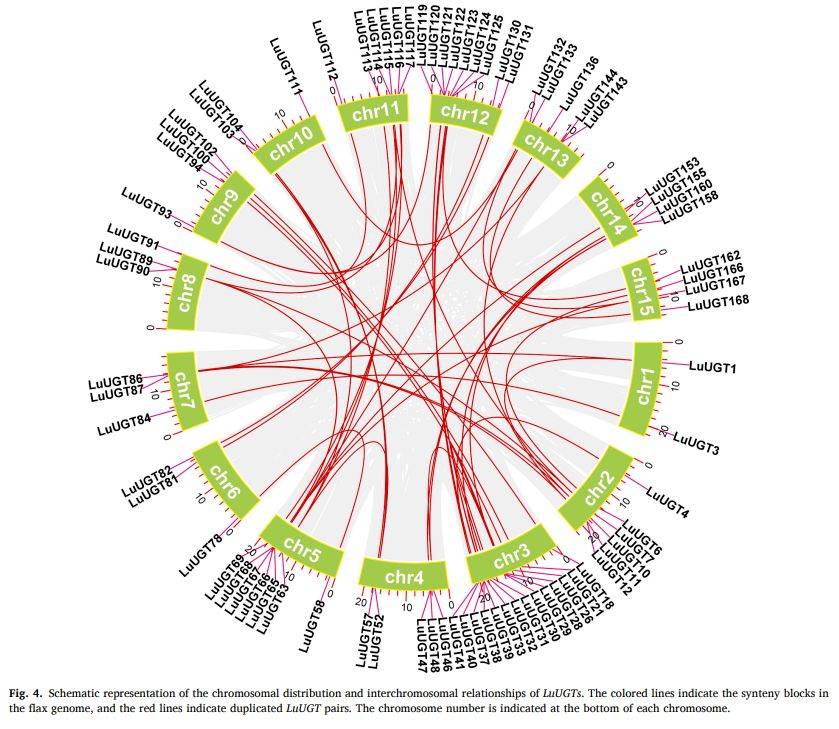
图例说明 :图4以circos形式展示15条染色体,红线标出46个LuUGT重复对,染色体编号位于外圈,彩色条带表示共线区块。

图5
段落内容 :为探讨LuUGT的跨物种进化,作者分别构建亚麻-拟南芥与亚麻-水稻共线性图。结果:23条LuUGT与18条AtUGT存在共线,而水稻仅3条LuUGT与3条OsUGT对应,显示该家族在双子叶间保守性远高于单子叶。图中灰色背景线为大片段共线区,红线突出UGT对,直观反映双子叶分化后UGT家族相对稳定,而单子叶与双子叶间剧烈分化,为选择拟南芥AtUGT72B1作为LuUGT175功能参照提供进化依据。
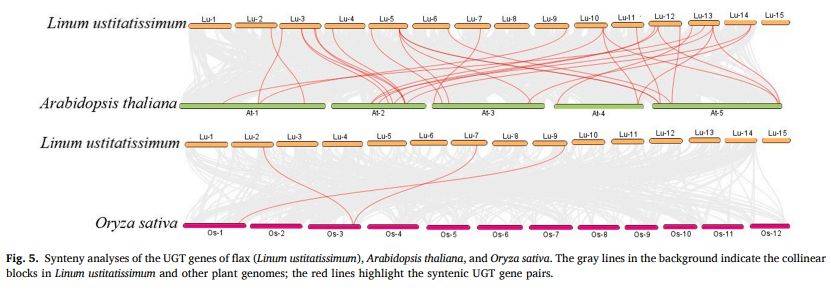
图例说明 :图5左侧为亚麻与拟南芥共线性,右侧为亚麻与水稻;红线标出直系同源UGT对,灰色背景表示染色体共线区块,染色体号标注于两端。
图6
段落内容 :基于NCBI-SRA公开RNA-seq数据,作者绘制190条LuUGT在根、茎、叶、花、纤维及种子5个发育阶段的表达热图。结果发现3条基因任何组织均不表达,83条呈组成型表达,64条表现组织特异;其中LuUGT175在茎与快速伸长期的纤维中FPKM最高,提示其可能参与纤维次生壁形成。Beanplot进一步展示表达分布,证实E组整体在茎部富集,为后续聚焦LuUGT175功能奠定表达证据。
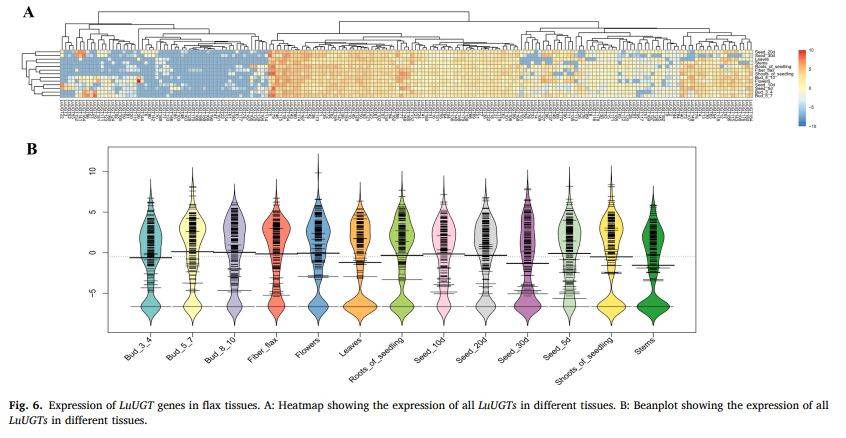
图例说明 :图6A为热图,颜色由蓝到红代表FPKM低-高;图6B为beanplot,横轴为组织,纵轴为log10(FPKM+1),曲线密度反映表达分布。

图7
段落内容 :CRISPR/Cas9敲除LuUGT175获得3个T0突变系(L4、L7、L8)。快速生长期(7周)茎徒手切片经Wiesner(盐酸-间苯三酚)与Mäule(高锰酸钾-盐酸-氨水)染色显示:野生型仅木质部呈淡紫红或棕色;突变体木质部颜色加深至暗紫红,且原本不染色的纤维层亦出现紫红/深棕,表明异位木质化。量化表明突变体茎 lignin 含量显著升高,证实LuUGT175缺失导致木质素过量积累。
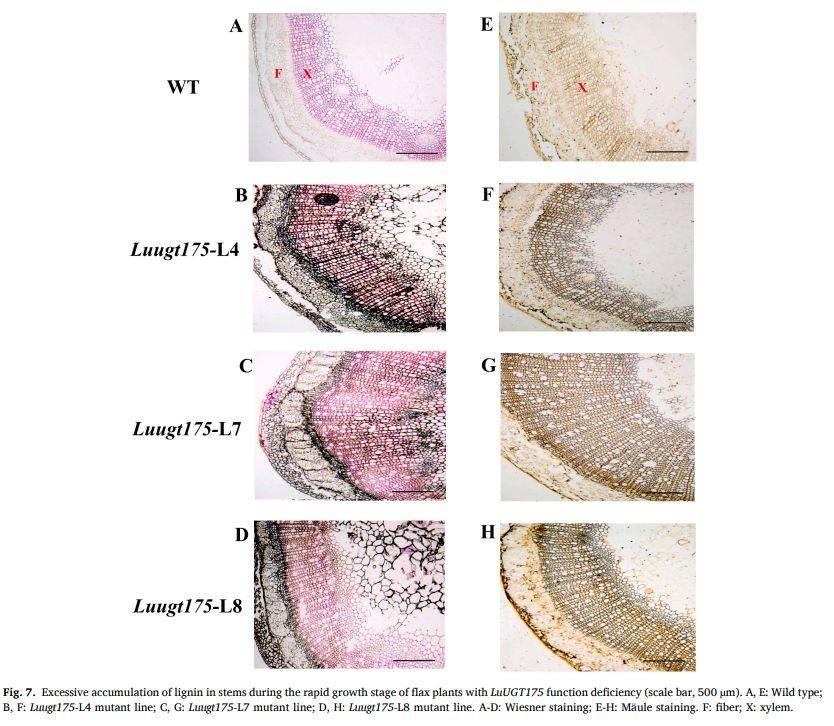
图例说明 :图7A-D为Wiesner染色,E-H为Mäule染色;标尺500 µm,F示纤维,X示木质部,突变体编号标注于图。

图8
段落内容 :为更清晰展示纤维束与木质部差异,作者提供图7局部放大(100 µm)。放大图可见野生型纤维细胞壁几乎不着色,突变体纤维壁呈明显紫红(Wiesner)或深棕(Mäule),且细胞间隙变小;木质部导管壁厚度亦增加。该结果在细胞水平直接证明LuUGT175通过糖基化调控木质素单体转运,其功能缺失使木质素在纤维细胞异位聚合,降低纤维柔韧性。
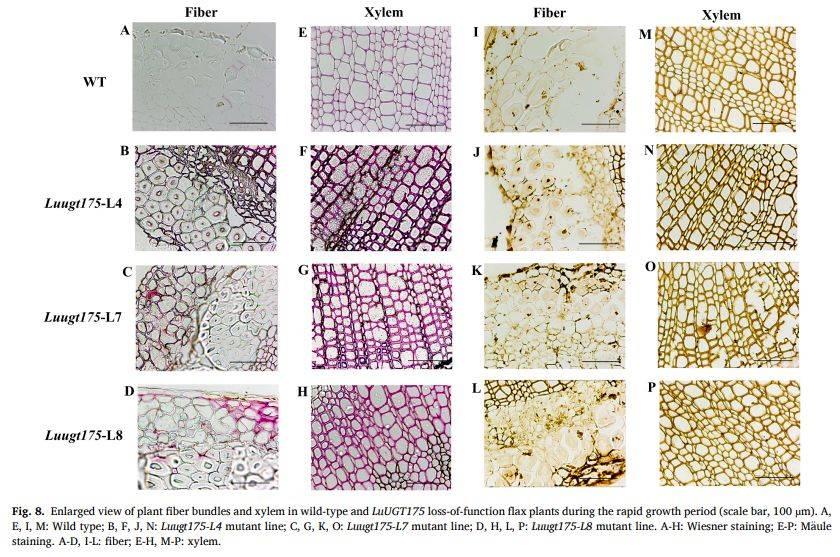
图例说明 :图8A-D、I-L为纤维束;E-H、M-P为木质部;左侧四行为Wiesner,右侧四行为Mäule;标尺100 µm,突变体系号标注于各图。

图9
段落内容 :作者对E组26条LuUGT与184条LuNAC转录因子进行表达相关性计算,发现17个LuNAC与20条LuUGT呈强负相关(r ≤ −0.6)。其中LuNAC61与LuUGT175相关系数达−0.86,位居负相关首位,提示其可能为上游负调控子。热图以蓝-白-红梯度展示相关系数,可直观比较不同NAC与UGT组合间的正负关系,为筛选LuNAC61作为候选调控基因提供数据依据。
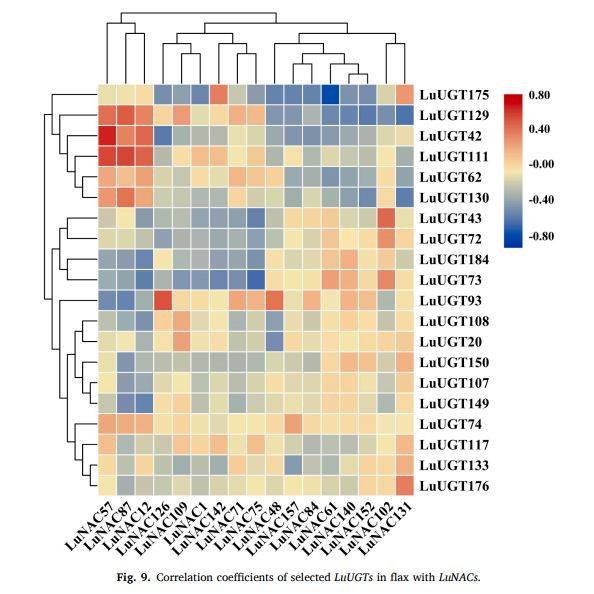
图例说明 :图9为横向热图,纵列为LuNAC编号,横行为LuUGT编号,颜色标尺由蓝(负相关)经白(0)至红(正相关),星号标出LuNAC61-LuUGT175对应格。

图10
段落内容 :基于共表达网络分析,LuUGT175与LuNAC61被聚于同一模块且边权重高,表明二者在转录水平存在显著关联。网络图以节点大小反映连接度,边线粗细代表共表达权重,LuNAC61处于模块核心位置,进一步支持其作为负调控枢纽。该结果与图9相关性分析互为印证,为后续酵母单杂、荧光素酶实验提供假设基础。
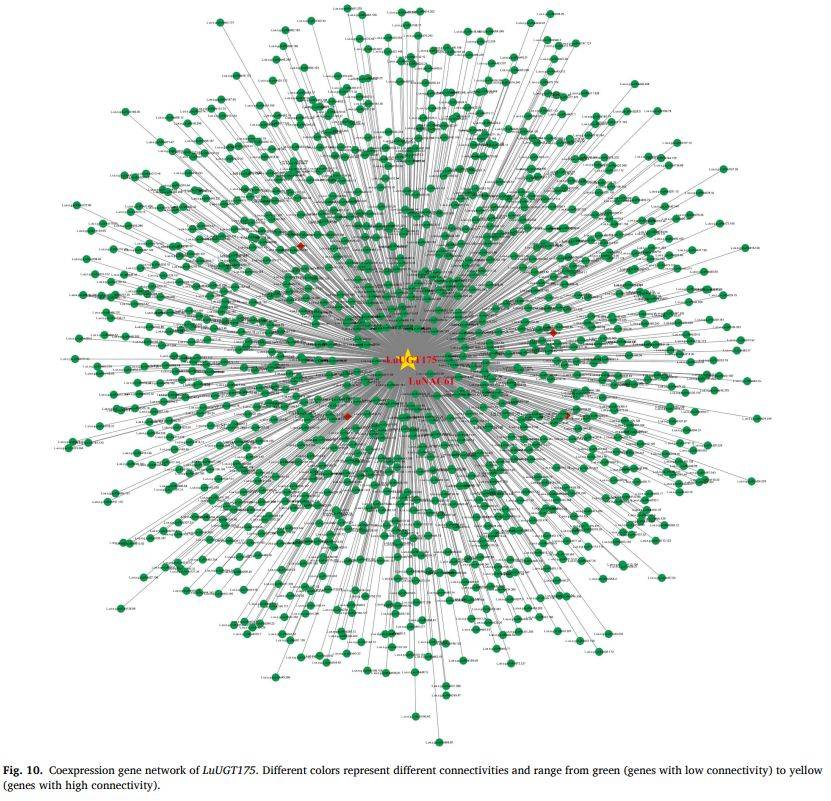
图例说明 :图10为无向网络图,绿色节点代表UGT,橙色节点代表NAC;边线越粗表示共表达权重越大,LuNAC61与LuUGT175用红色高亮。

图11
段落内容 :作者通过烟草瞬时表达证实LuUGT175-GFP融合蛋白定位于细胞质与细胞核,符合其作为糖基转移酶参与胞质糖基化及核内信号调控的双重角色。酵母单杂交显示LuNAC61能驱动报告基因在150 ng/ml AbA选择性培养基生长,而空载体对照不能,表明其可直接结合LuUGT175启动子。双荧光素酶实验进一步在植物体内验证:LuNAC61显著抑制LuUGT175启动子活性,荧光信号低于对照50%以上,综合证明LuNAC61为LuUGT175上游负调控因子。
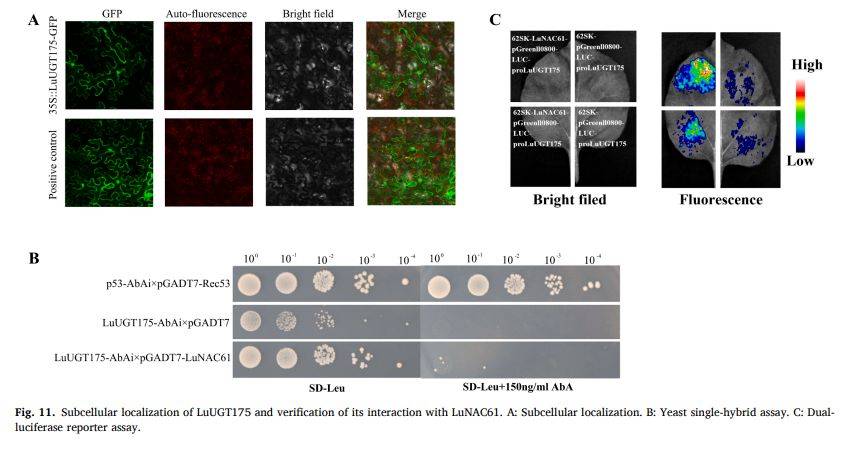
图例说明 :图11A为共聚焦GFP亚细胞定位,标尺50 µm;B为酵母单杂平板,左-右依次为负对照、正对照、实验组;C为烟草叶片荧光成像,上排为对照,下排为LuNAC61效应组,亮度差异直观展示抑制效果。
四、总结此文
基因组挖掘:以PSPG盒为标签,系统鉴定190个LuUGTs,分布于15条染色体,分属16个进化组(A–P),其中E组与拟南芥AtUGT72B1直系同源。
进化与复制:全基因组重复与片段重复是家族扩张主因;68.4%成员含内含子,保守基序1–3在蛋白C端构成糖基转移酶催化核心。
表达谱:RNA-seq显示187个LuUGTs在至少一组织中表达,83个组成型表达,64个呈组织特异;LuUGT175在茎与纤维优势高表达。
功能验证:CRISPR敲除LuUGT175获得3个T0突变系,植株矮化、茎木质部与纤维层出现异位木质化,Wiesner/Mäule染色显著加深,木质素含量升高。
调控机制:共表达与相关性分析筛选到负相关转录因子LuNAC61(r = −0.86);酵母单杂、双荧光素酶实验证实LuNAC61可直接结合并抑制LuUGT175启动子活性。
意义:首次阐明亚麻UGT家族全貌,揭示LuUGT175-糖基化-木质素通路,为利用生物技术降低纤维木质素、改善可纺性提供关键靶点。
五、生成思维导图(文字版)
基因组鉴定
190 LuUGTs │ 分16组 │ 染色体不均分布 │ 全基因组/片段重复扩张
表达特征
187个表达 │ 83组成型 │ 64组织特异 │ LuUGT175茎/纤维高表达
功能验证
CRISPR敲除 → 矮化 │ 茎木质部染色加深 │ 纤维异位木质化 │ 木质素含量↑
调控网络
共表达筛选 │ LuNAC61负相关(r=−0.86) │ 酵母单杂+双荧光素酶验证直接结合
应用前景
低木质素分子育种 │ 改善纤维柔软度与可纺性 │ 减少脱胶污染







评论